Пептидите се класа на соединенија формирани со поврзување на повеќе амино киселини преку пептидни врски.Тие се сеприсутни во живите организми.До сега, десетици илјади пептиди се пронајдени во живите организми.Пептидите играат важна улога во регулирањето на функционалните активности на различни системи, органи, ткива и клетки и во животните активности и често се користат во функционална анализа, истражување на антитела, развој на лекови и други полиња.Со развојот на биотехнологијата и технологијата за синтеза на пептиди, се повеќе и повеќе пептидни лекови се развиваат и се применуваат во клиниката.
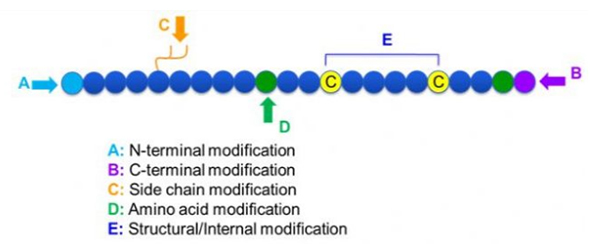
Има широк спектар на модификации на пептиди, кои едноставно можат да се поделат на пост-модификација и модификација на процесот (со користење на изведена модификација на аминокиселини), и модификација на N-терминалот, модификација на C-терминалниот, модификација на страничниот синџир, модификација на аминокиселина, модификација на скелетот, итн., во зависност од местото на модификација (слика 1).Како важно средство за промена на структурата на главниот синџир или групите на страничните синџири на пептидните синџири, модификацијата на пептидот може ефикасно да ги промени физичките и хемиските својства на пептидните соединенија, да ја зголеми растворливоста во вода, да го продолжи времето на дејство in vivo, да ја промени нивната биолошка дистрибуција, да ја елиминира имуногеноста. , намалување на токсичните несакани ефекти итн. Во овој труд се воведени неколку главни стратегии за модификација на пептиди и нивните карактеристики.
1. Циклизација
Цикличните пептиди имаат многу примени во биомедицината, а многу природни пептиди со биолошка активност се циклични пептиди.Бидејќи цикличните пептиди имаат тенденција да бидат поригидни од линеарните пептиди, тие се исклучително отпорни на дигестивниот систем, можат да преживеат во дигестивниот тракт и покажуваат посилен афинитет за целните рецептори.Циклизацијата е најдиректниот начин за синтеза на циклични пептиди, особено за пептиди со голем структурен скелет.Според режимот на циклизација, може да се подели на тип на синџир од страничен синџир, тип на страничен синџир, терминал - тип на терминал (тип од крај до крај).
(1) страничен синџир од страничен синџир
Најчестиот тип на циклизација од страничен до страничен синџир е дисулфидното премостување помеѓу остатоците од цистеин.Оваа циклизација е воведена со пар остатоци од цистеин кои се дезаштитени и потоа оксидирани за да формираат дисулфидни врски.Полицикличната синтеза може да се постигне со селективно отстранување на заштитните групи на сулфидрил.Циклизацијата може да се направи или во растворувач по дисоцијација или на смола пред дисоцијација.Циклизацијата на смоли може да биде помалку ефикасна од циклизацијата на растворувачите бидејќи пептидите на смоли не формираат лесно циклифицирани конформации.Друг тип на страничен синџир - цикализација на страничниот синџир е формирање на амидна структура помеѓу остаток од аспарагинска киселина или глутаминска киселина и базната амино киселина, што бара заштитната група на страничниот синџир да може селективно да се отстрани од полипептидот или на смолата или по дисоцијација.Третиот тип на страничен синџир - циклизација на страничен синџир е формирање на дифенил етери од тирозин или р-хидроксифенилглицин.Овој тип на циклизација кај природните производи се среќава само во микробните производи, а производите за циклизација често имаат потенцијална медицинска вредност.Подготовката на овие соединенија бара уникатни услови за реакција, така што тие често не се користат во синтезата на конвенционалните пептиди.
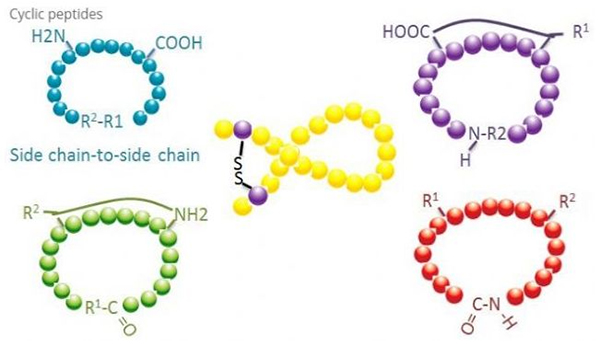
(2) од терминал до страничен синџир
Цикализацијата на синџирот од крајната страна обично го вклучува C-терминалот со амино групата на страничниот синџир на лизин или орнитин, или N-терминалот со страничниот ланец на аспарагинската киселина или глутаминската киселина.Друга цикализација на полипептид е направена со формирање на етерски врски помеѓу терминалот C и страничните синџири на серин или треонин.
(3) Терминален или тип од глава до опашка
Полипептидите на синџирот може или да се циклираат во растворувач или да се фиксираат на смола со циклирање на страничен синџир.Ниски концентрации на пептиди треба да се користат при централизација на растворувачи за да се избегне олигомеризација на пептидите.Приносот на синтетички прстенести полипептид од глава до опашка зависи од низата на полипептидот на синџирот.Затоа, пред да се подготват циклични пептиди во голем обем, прво треба да се создаде библиотека на можни оловни пептиди со синџир, по што ќе следи циклизација за да се најде низата со најдобри резултати.
2. N-метилација
N-метилацијата првично се јавува во природните пептиди и се воведува во синтезата на пептидите за да се спречи формирањето на водородни врски, со што пептидите се поотпорни на биоразградување и клиренс.Синтезата на пептиди користејќи деривати на N-метилирани аминокиселини е најважниот метод.Дополнително, Mitsunobu реакцијата на N-(2-нитробензен сулфонил хлорид) полипептид-смола посредници со метанол, исто така, може да се користи.Овој метод се користи за подготовка на библиотеки на циклични пептиди кои содржат N-метилирани амино киселини.
3. Фосфорилација
Фосфорилацијата е една од најчестите пост-преведувачки модификации во природата.Во човечките клетки, повеќе од 30% од протеините се фосфорилирани.Фосфорилацијата, особено реверзибилната фосфорилација, игра важна улога во контролирањето на многу клеточни процеси, како што се трансдукцијата на сигналот, генската експресија, регулацијата на клеточниот циклус и цитоскелетот и апоптозата.
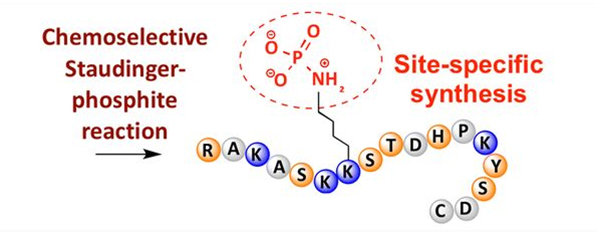
Фосфорилацијата може да се забележи кај различни остатоци од аминокиселини, но најчести цели за фосфорилација се остатоците од серин, треонин и тирозин.Дериватите на фосфотирозин, фосфотреонин и фосфосерин можат или да се внесат во пептидите за време на синтезата или да се формираат по синтезата на пептидите.Селективна фосфорилација може да се постигне со користење на остатоци од серин, треонин и тирозин кои селективно ги отстрануваат заштитните групи.Некои реагенси за фосфорилација, исто така, можат да внесат групи на фосфорна киселина во полипептидот со пост-модификација.Во последниве години, фосфорилација на лизин специфична за локацијата е постигната со користење на хемиски селективна Стаудингер-фосфитна реакција (Слика 3).
4. Миристоилација и палмитоилација
Ацилирањето на N-терминалот со масни киселини им овозможува на пептидите или протеините да се врзат за клеточните мембрани.Миридамоилираната секвенца на N-терминалот овозможува протеинските кинази од семејството Src и протеините на реверзна транскриптаза Gaq да бидат насочени да се врзат за клеточните мембрани.Миристичната киселина беше поврзана со N-терминалот на смола-полипептидот користејќи стандардни реакции на спојување, а добиениот липопептид може да се дисоцира во стандардни услови и да се прочисти со RP-HPLC.
5. Гликозилација
Гликопептидите како ванкомицин и теиколанин се важни антибиотици за третман на бактериски инфекции отпорни на лекови, а други гликопептиди често се користат за стимулирање на имунолошкиот систем.Дополнително, бидејќи многу микробни антигени се гликолизирани, од големо значење е да се проучуваат гликопептидите за подобрување на терапевтскиот ефект на инфекцијата.Од друга страна, откриено е дека протеините на клеточната мембрана на клетките на туморот покажуваат абнормална гликозилација, што ги прави гликопептидите да играат важна улога во истражувањето на имунолошката одбрана на ракот и туморот.Гликопептидите се подготвуваат со методот Fmoc/t-Bu.Гликолизираните остатоци, како што се треонин и серин, често се внесуваат во полипептидите со fMOC активирани со пентафлуорофенол естер за да се заштитат гликолизираните амино киселини.
6. Изопрен
Изопентадиенилацијата се јавува на остатоците од цистеин во страничниот ланец во близина на C-терминалот.Протеинскиот изопрен може да го подобри афинитетот на клеточната мембрана и да формира протеинско-протеинска интеракција.Изопентадиенираните протеини вклучуваат тирозин фосфатаза, мала GTase, молекули на кокаперон, нуклеарна ламина и центромерни врзувачки протеини.Изопренските полипептиди може да се подготват со користење на изопрен на смоли или со воведување деривати на цистеин.
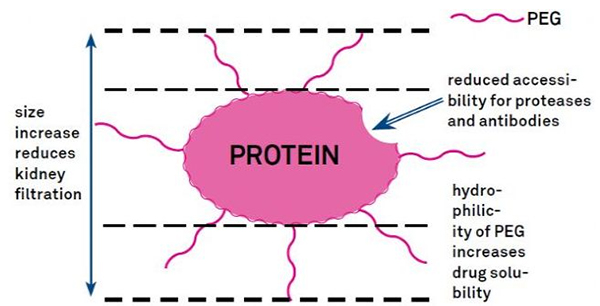
7. Модификација на полиетилен гликол (PEG).
Модификацијата на PEG може да се користи за подобрување на протеинската хидролитичка стабилност, биодистрибуција и растворливост на пептиди.Воведувањето на PEG синџири во пептидите може да ги подобри нивните фармаколошки својства и исто така да ја инхибира хидролизата на пептидите со протеолитички ензими.PEG пептидите минуваат низ гломеруларниот капиларен пресек полесно од обичните пептиди, што значително го намалува реналниот клиренс.Поради продолжениот активен полуживот на PEG пептидите in vivo, нормалното ниво на третман може да се одржи со помали дози и поретко пептидни лекови.Сепак, модификацијата на PEG има и негативни ефекти.Големи количини на PEG го спречуваат ензимот да го деградира пептидот и исто така го намалуваат врзувањето на пептидот со целниот рецептор.Но, нискиот афинитет на PEG пептидите обично се компензира со нивниот подолг фармакокинетски полуживот, а со тоа што се присутни во телото подолго, PEG пептидите имаат поголема веројатност да се апсорбираат во целните ткива.Затоа, спецификациите на PEG полимерот треба да се оптимизираат за оптимални резултати.Од друга страна, PEG пептидите се акумулираат во црниот дроб поради намален ренален клиренс, што резултира со макромолекуларен синдром.Затоа, модификациите на PEG треба да се дизајнираат повнимателно кога пептидите се користат за тестирање на лекови.
Вообичаените модификациски групи на PEG модификатори грубо може да се сумираат на следниов начин: амино (-амин) -NH2, аминометил-Ch2-NH2, хидрокси-OH, карбокси-Cooh, сулфхидрил (-тиол) -SH, Малеимид -MAL, сукцинимид карбонат - SC, сукцинимид ацетат -SCM, сукцинимид пропионат -SPA, n-хидроксисуцинимид -NHS, акрилат-ch2ch2cooh, алдехид -CHO (како што е пропион-алд, бутираЛД), акрилна база (-акрилат-акрл), азидо-инил-азид, Биотин, флуоресцеин, глутарил -GA, акрилат хидразид, алкин-алкин, р-толуенсулфонат -OTs, сукцинимид сукцинат -SS, итн. Дериватите на PEG со карбоксилни киселини може да се спојат со n-терминални амини или странични синџири на лизин.Амино-активираниот PEG може да се спои со страничните синџири на аспарагинска киселина или глутаминска киселина.Неправилно активираниот PEG може да се конјугира со меркаптан од целосно дезаштитени странични синџири на цистеин [11].PEG модификаторите обично се класифицираат на следниов начин (забелешка: mPEG е метокси-PEG, CH3O-(CH2CH2O)n-CH2CH2-OH):
(1) PEG модификатор со директен ланец
mPEG-SC, mPEG-SCM, mPEG-SPA, mPEG-OTs, mPEG-SH, mPEG-ALD, mPEG-butyrALD, mPEG-SS
(2) бифункционален PEG модификатор
HCOO-PEG-COOH, NH2-PEG-NH2, OH-PEG-COOH, OH-PEG-NH2, HCl·NH2-PEG-COOH, MAL-PEG-NHS
(3) разгранувачки PEG модификатор
(mPEG)2-NHS, (mPEG)2-ALD, (mPEG)2-NH2, (mPEG)2-MAL
8. Биотинизација
Биотинот може силно да се врзува со авидин или стрептавидин, а силата на врзување е дури блиску до ковалентна врска.Пептидите означени со биотин најчесто се користат во имуноанализа, хистоцитохемија и проточна цитометрија базирана на флуоресценција.Етикетирани антибиотински антитела, исто така, може да се користат за врзување на биотинилирани пептиди.Етикетите на биотин често се прикачени на страничниот синџир на лизин или на терминалот N.6-аминокапроична киселина често се користи како врска помеѓу пептидите и биотинот.Врската е флексибилна во врзувањето за подлогата и подобро се врзува во присуство на стерична пречка.
9. Флуоресцентно означување
Флуоресцентното означување може да се користи за следење на полипептидите во живите клетки и за проучување на ензимите и механизмите на дејство.Триптофанот (Trp) е флуоресцентен, така што може да се користи за внатрешно означување.Емисиониот спектар на триптофан зависи од периферната средина и се намалува со намалување на поларитетот на растворувачите, својство кое е корисно за откривање на структурата на пептидот и врзувањето на рецепторот.Флуоресценцијата на триптофан може да се угасне со протонирана аспарагинска киселина и глутаминска киселина, што може да ја ограничи неговата употреба.Групата Dansyl хлорид (Dansyl) е високо флуоресцентна кога се врзува за амино група и често се користи како флуоресцентна ознака за амино киселини или протеини.
Флуоресцентна резонанца Конверзијата на енергија (FRET) е корисна за ензимски студии.Кога се применува FRET, полипептидот на подлогата обично содржи група за означување на флуоресценција и група за гаснење на флуоресценција.Обележените флуоресцентни групи се гасат од гаснечот преку пренос на енергија без фотон.Кога пептидот е дисоциран од ензимот за кој станува збор, групата за означување емитува флуоресценција.
10. Полипептиди во кафез
Пептидите во кафез имаат оптички отстранливи заштитни групи кои го штитат пептидот од врзување со рецепторот.Кога е изложен на УВ зрачење, пептидот се активира, враќајќи го неговиот афинитет кон рецепторот.Бидејќи ова оптичко активирање може да се контролира според времето, амплитудата или локацијата, пептидите во кафез може да се користат за проучување на реакциите што се случуваат во клетките.Најчесто користени заштитни групи за полипептиди во кафез се 2-нитробензилните групи и нивните деривати, кои можат да се воведат во синтезата на пептидите преку заштитни деривати на амино киселини.Деривати на амино киселини кои се развиени се лизин, цистеин, серин и тирозин.Дериватите на аспартат и глутамат, сепак, не се користат вообичаено поради нивната подложност на цикализација за време на синтезата и дисоцијацијата на пептидите.
11. Полиантигенски пептид (MAP)
Кратките пептиди обично не се имуни и мора да се спојат со протеините-носители за да се произведат антитела.Полиантигенскиот пептид (MAP) е составен од повеќе идентични пептиди поврзани со јадра на лизин, кои можат конкретно да изразат имуногени со висока моќност и може да се користат за подготовка на парцели на протеински носачи на пептиди.MAP полипептидите може да се синтетизираат со синтеза на цврста фаза на MAP смола.Сепак, нецелосното спојување резултира со исчезнати или скратени пептидни синџири на некои гранки и на тој начин не ги покажува својствата на оригиналниот MAP полипептид.Како алтернатива, пептидите може да се подготват и прочистат одделно и потоа да се спојат со MAP.Пептидната низа прикачена на јадрото на пептидот е добро дефинирана и лесно се карактеризира со масена спектрометрија.
Заклучок
Модификацијата на пептидите е важно средство за дизајнирање на пептиди.Хемиски модифицираните пептиди не само што можат да одржат висока биолошка активност, туку и ефикасно да ги избегнат недостатоците на имуногеноста и токсичноста.Во исто време, хемиската модификација може да им даде на пептидите некои нови одлични својства.Во последниве години, методот на активирање на CH за пост-модификација на полипептидите е брзо развиен и постигнати се многу важни резултати.
Време на објавување: Мар-20-2023